Petri Rautiala has joined us on a Newton International Fellowship, to develop theory on the epigenetical evolution of social behaviour. Welcome to the group, Petri!
[New Paper] Sexual selection modulates genetic conflicts and patterns of genomic imprinting
Faria GS, Varela SAM & Gardner A (in press) Sexual selection modulates genetic conflicts and patterns of genomic imprinting. Evolution doi: 10.1111/evo.13153
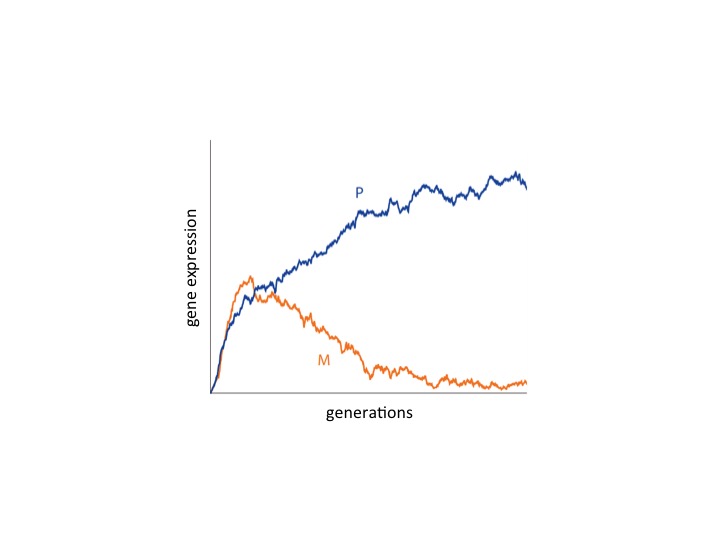
Recent years have seen a surge of interest in linking the theories of kin selection and sexual selection. In particular, there is a growing appreciation that kin selection, arising through demographic factors such as sex–biased dispersal, may modulate sexual conflicts, including in the context of male-female arms races characterized by coevolutionary cycles. However, evolutionary conflicts of interest need not only occur between individuals, but may also occur within individuals, and sex-specific demography is known to foment such intragenomic conflict in relation to social behavior. Whether and how this logic holds in the context of sexual conflict – and, in particular, in relation to coevolutionary cycles – remains obscure. We develop a kin-selection model to investigate the interests of different genes involved in sexual and intragenomic conflict, and we show that consideration of these conflicting interests yields novel predictions concerning parent–of–origin-specific patterns of gene expression and the detrimental effects of different classes of mutation and epimutation at loci underpinning sexually–selected phenotypes.
PhD Opportunity: Adaptation of genes, individuals and societies

I am offering a PhD studentship on “Theory of social evolution: adaptation of genes, individuals and societies” (closing date Mon 5 Dec 2016, with a flexible start date of around Sep 2017). More details can be found here.
[New Group Members] Bekah Best & Mauricio González Forero
Two new people have joined our research group: Bekah Best will be doing her undergraduate project on genomic imprinting, and Mauricio González Forero has brought a Marie Skłodowska-Curie Fellowship to St Andrews to develop theory on brain evolution. Welcome to the group, Bekah and Mauricio!
[New Paper] The strategic revolution
Gardner A (2016) The strategic revolution. Cell 166, 1345-1348.

On the 40th anniversary of the publication of Richard Dawkins’s The Selfish Gene, we explore the origins of cynical, strategic thinking in evolutionary biology, investigate how this illuminated the sexual and social lives of animals, and assess Dawkins’s suggestion that evolution is best understood by taking the gene’s-eye view.
[New Paper] Ecology of sex explains patterns of helping in arthropod societies
Davies NG, Ross L & Gardner A (2016) The ecology of sex explains patterns of helping in arthropod societies. Ecology Letters doi: 10.1111/ele.12621
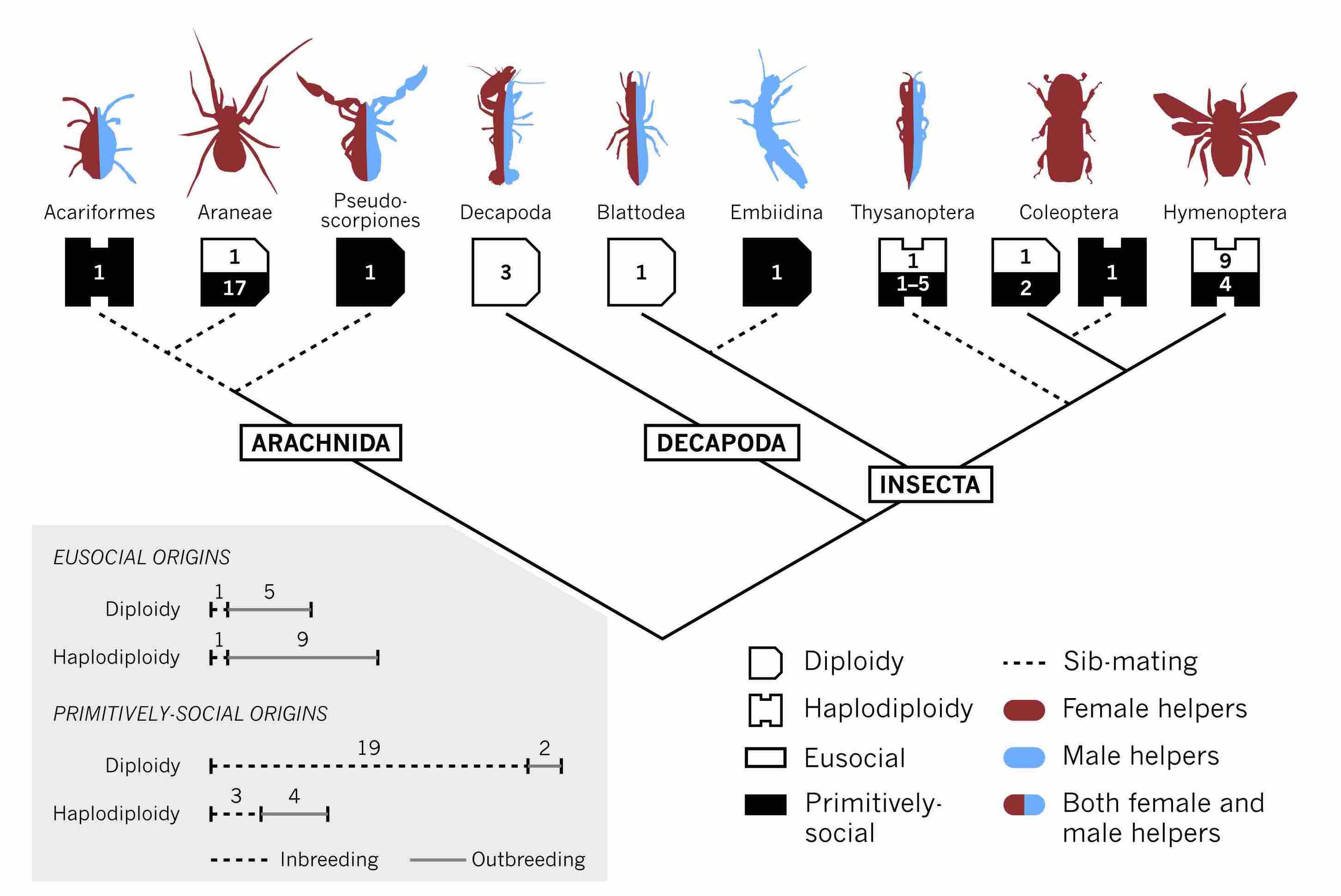
Across arthropod societies, sib-rearing (e.g. nursing or nest defence) may be provided by females, by males or by both sexes. According to Hamilton’s ‘haplodiploidy hypothesis’, this diversity reflects the relatedness consequences of diploid vs. haplodiploid inheritance. However, an alternative ‘preadaptation hypothesis’ instead emphasises an interplay of ecology and the co-option of ancestral, sexually dimorphic traits for sib-rearing. The preadaptation hypothesis has recently received empirical support, but remains to be formalised. Here, we mathematically model the coevolution of sex-specific helping and sex allocation, contrasting these hypotheses. We find that ploidy per se has little effect. Rather, the ecology of sex shapes patterns of helping: sex-specific preadaptation strongly influences who helps; a freely adjustable sex ratio magnifies sex biases and promotes helping; and sib-mating, promiscuity, and reproductive autonomy also modulate the sex and abundance of helpers. An empirical survey reveals that patterns of sex-specific helping in arthropod taxa are consistent with the preadaptation hypothesis.
[New Paper] Misconceptions on the application of biological market theory to the mycorrhizal symbiosis
Kiers ET, West SA, Wyatt GAK, Gardner A, Bücking H & Werner G (2016) Misconceptions on the application of biological market theory to the mycorrhizal symbiosis. Nature Plants 2, 16063.
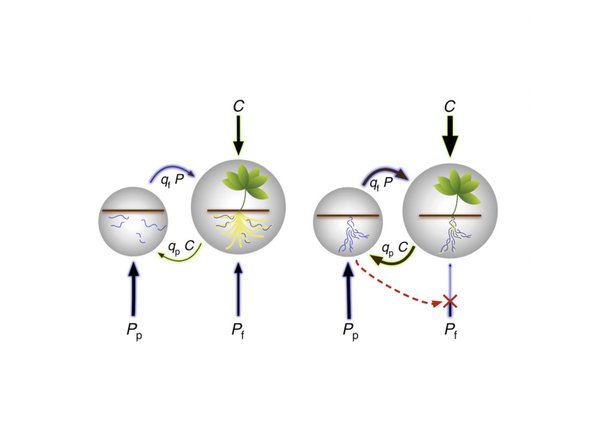
The symbiosis between plants and arbuscular mycorrhizal fungi has been described as a biological market based on evidence that plants supply more carbohydrates to fungal partners that provide more soil nutrients, and vice versa. A recent paper by Walder and van der Heijden challenges this view. However, their challenge is based on misunderstandings of biological market theory, and evolutionary theory more generally.
[New Paper] Sex ratios, virginity & local resource enhancement in a quasisocial parasitoid
Kapranas A, Hardy ICW, Tang X, Gardner A & Li B (2016) Sex ratios, virginity and local resource enhancement in a quasisocial parasitoid. Entomologia Experimentalis et Applicata 159, 243-251.

Sclerodermus harmandi (Buysson) (Hymenoptera: Bethylidae) is an economically beneficial species of parasitoid wasp that has an unusual level of sociality: groups of female foundresses reproduce on a single host and exhibit cooperative post-ovipositional brood care. The beneficial effects females have on each other’s reproductive success provide, via the theory of local resource enhancement (LRE), an explanation for their female-biased progeny sex ratios, which is part of the same framework for understanding sex-ratio evolution as the more often invoked theory of local mate competition (LMC). Here, we show that S. harmandi sex ratios are overdispersed, with high variance largely attributable to the common occurrence (60%) of developmental mortality. Developmental mortality is also positively associated with the proportion of broods which contain only females at emergence (virgin broods). Virginity is more common when broods are produced by fewer foundresses. Virginity is expected to be disadvantageous under LRE, as it is under LMC, but theory for LRE is less extensively developed. We suggest approaches for the development of LRE theory, in particular using models of ‘population elasticity’ in which the intensity of kin competition is reduced because extra resources are available to local populations that are more cooperative. For S. harmandi, such extra resources may include large hosts that can only be successfully utilised if multiple foundresses cooperate.
[New Paper] Intragenomic conflict in polyembryonic parasitoid wasps
Rautiala P & Gardner A (in press) Intragenomic conflict over soldier allocation in polyembryonic parasitoid wasps. American Naturalist. doi: 10.1086/685082
Understanding the selection pressures that have driven the evolution of sterile insect castes has been the focus of decades of intense scientific debate. An amenable empirical test bed for theory on this topic is provided by the sterile-soldier caste of polyembryonic parasitoid wasps. The function of these soldiers has been a source of controversy, with two basic hypotheses emerging: the “brood-benefit” hypothesis that they provide an overall benefit for their siblings and the “sex-ratio-conflict” hypothesis that the soldiers mediate a conflict between brothers and sisters by killing their opposite-sex siblings. Here, we investigate the divergent sex-ratio optima of a female embryo’s maternal-origin and paternal-origin genes, to determine the potential for, and direction of, intragenomic conflict over soldiering. We then derive contrasting empirically testable predictions concerning the patterns of genomic imprinting that are expected to arise out of this intragenomic conflict, for the brood-benefit versus the sex-ratio-conflict hypothesis of soldier function.
[New Paper] Restricting mutualistic partners to enforce trade reliance
Wyatt GAK, Kiers ET, Gardner A & West SA (2016) Restricting mutualistic partners to enforce trade reliance. Nature Communications 7, 10322 doi: 10.1038/ncomms10322
Mutualisms are cooperative interactions between members of different species, often involving the trade of resources. Here, we suggest that otherwise-cooperative mutualists might be able to gain a benefit from actively restricting their partners’ ability to obtain resources directly, hampering the ability of the restricted partner to survive and/or reproduce without the help of the restricting mutualist. We show that (i) restriction can be favoured when it makes the resources of the restricting individual more valuable to their partner, and thus allows them to receive more favourable terms of trade; (ii) restriction maintains cooperation in conditions where cooperative behaviour would otherwise collapse; and (iii) restriction can lead to either an increase or decrease in a restricted individual’s fitness. We discuss the applicability of this scenario to mutualisms such as those between plants and mycorrhizal fungi. These results identify a novel conflict in mutualisms as well as several public goods dilemmas, but also demonstrate how conflict can help maintain cooperation.